全文PDFは以下からダウンロードできます。
JaSMInJapan Registration System for Metabolic & Inherited Diseases


先天性ケトン体代謝異常症の話
岐阜大学大学院医学系研究科 小児病態学 深尾 敏幸
1. ケトン体とは?
ケトン体とはアセト酢酸と3ヒドロキシ酪酸という炭素が4つのカルボン酸(COOHがある酸)です。皆さんがよくご存知の酢酸(酢)は炭素が2つのカルボン酸です。ケトン体は酢酸よりは酸性度は強くないですが、それでも強い酸です。ですからケトン体が蓄積すればやっぱり体は酸性に傾きます(アシドーシスといいます)。有機酸といわれるものも炭素数が3~6個ぐらいのカルボン酸で、ケトン体も有機酸の1つと言えます。乳酸とかピルビン酸というのも聞いたことがあるかもしれません、これは炭素が3つの有機酸になります。
2. ケトン体の役割
ケトン体はどんな役目があるのでしょうか?子供が熱を出したり、胃腸風邪で嘔吐をして食欲がない時などに、尿ケトンが簡単に陽性になります。発熱や食欲不振時にケトンが陽性になるのでケトン体は悪者のように言われますが、違います。ケトン体は大切なエネルギー源として肝臓で作られます。肝臓は脂肪に蓄えていたエネルギーをケトン体という形に変えて肝臓以外の臓器、特に脳に提供してくれているのです。この働きによってヒト(特に子供)では低血糖になることを防いでいるのです。ですから、脂肪の分解ができない脂肪酸代謝異常症(極長鎖アシル-CoA脱水素酵素欠損症やカルニチンパルミトイルトランスフェラーゼ欠損症など)では、このエネルギー源であるケトン体ができないために、グルコースが消費され低血糖を起こしてしまいます。
では、どれくらい空腹になると危険なのでしょうか?一般的には発熱とかストレスがない状況では、食事由来のグルコースで食後3~4時間は血糖が保てます。ですから、3時間ごとに母乳ミルクを飲んでいる赤ちゃんではケトン体産生が亢進することはありません。4時間以上経つと血糖を保つために肝臓に貯めてあったグリコーゲンを分解して血糖を保ちます。これが少なくなってくるといよいよ血糖維持は、糖新生系と脂肪酸酸化系―ケトン体産生系にかかってきます。このグルコーゲンからのスイッチは年齢によって違いますし、発熱やストレス状態があるかどうかでも違います。しかし、小児では8~16時間ぐらいで糖新生系と脂肪酸酸化系―ケトン体産生系に血糖維持機構が移ってきます。ですから、ケトン体代謝異常症ではこの時期になって症状が出てくることになります。先ほどから繰り返していますが、発熱や胃腸炎などのストレス時には状態の良い時より早くこのスイッチがおきますので注意が必要です。発作を防ぐにはそれまでの時間に十分なグルコースを取ることが重要となります。発作を防ぐことが発作を治療するより重要です。
3. ケトン体代謝異常症
ケトン体代謝を図1に示します。
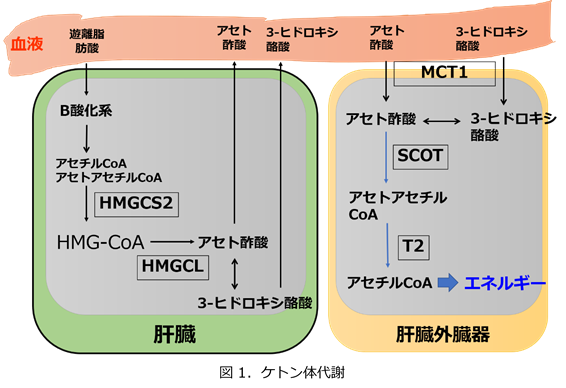
(1)ケトン体産生障害
広い意味で脂肪酸ベータ酸化系の異常症は、ケトン体産生の異常となります。ただし、脂肪酸のベータ酸化系異常症は、肝臓におけるケトン体産生異常にプラスして、心臓と筋肉における脂肪酸のベータ酸化の異常も伴うため、心筋症や横紋筋融解(筋肉痛、CK上昇)を起こします。ケトン体産生障害の臨床像は、通常では耐えられる程度の空腹、ストレスでケトン体がうまく産生されないために、低血糖になります。食事をしていればケトン体に頼らなくてもいいので低血糖は起きません。脂肪酸のベータ酸化系異常症以外にミトコンドリアHMG-CoA合成酵素(図ではHMGCS2)欠損症、HMG-CoAリアーゼ(図ではHMGCL)欠損症があります。少し細かいことを言いますとミトコンドリアHMG-CoA合成酵素欠損症ではロイシンからのHMG-CoAは作られるため、ロイシン由来のケトン体は産生できます。しかし、HMG-CoAリアーゼ欠損症ではロイシンからのケトン体の産生もできず、またロイシンからの代謝産物の蓄積が起きるため、ミトコンドリアHMG-CoA合成酵素欠損症に比べると重症と言えます。
(2)ケトン体利用障害
ケトン体は肝臓で低血糖を防ごうとして産生されるのに、肝外臓器がこれを利用できない疾患です。このため、空腹とかストレス状態で血中ケトンが蓄積して低血糖になるよりも前にアシドーシスを起こします。ケトン体利用障害で低血糖がある場合は著しいアシドーシスとなるはずです。利用障害をきたす疾患には、モノカルボン酸トランスポーター1(図ではMCT1)欠損症とサクシニル-CoA:3-ケト酸CoAトランスフェラーゼ(図ではSCOT)欠損症(スクシニル….と記載することもあります)、そしてイソロイシン代謝にも関係するβ-ケトチオラーゼ(図ではT2)欠損症が挙げられます。これらの疾患も原則的には、食事をしてケトン体の産生を必要としない状況では問題はなく無症状と言えます。
4. ケトン体代謝異常症と新生児マススクリーニング
ケトン体代謝異常症の中で新生児マススクリーニングの1次対象疾患となっているのはHMG-CoAリアーゼ欠損症ですが、タンデムマスパイロット研究と2014年からの公的スクリーニングで未だ症例は見つかっておらず、日本における疾患頻度は非常に稀と考えられています。β-ケトチオラーゼ欠損症は2次対象疾患ですが、2014年に公的スクリーニングが開始されて2例が確定診断されています。そのほかの疾患はマススクリーニング対象疾患となっていません。β-ケトチオラーゼ欠損症も新生児マススクリーニングでは見逃す可能性のある疾患であり、新生児マススクリーニングで問題なかったからといってケトン体代謝異常症ではないとは決して言えません。
5. ケトン体の産生系は進化の過程で形成された
ここからは少し臨床を離れた話をします。生物系に興味のある方は読んでみてください。
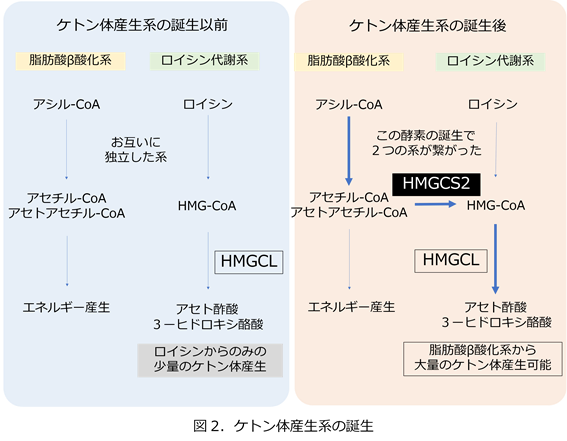
もともと生物にはケトン体代謝というのはなかったようです(図2)。肝臓には脂肪酸のベータ酸化系とロイシンというアミノ酸の分解系は存在していました。ロイシンの分解系にHMG-CoAリアーゼ(図ではHMGCL)という酵素は存在していました。ですから、肝臓で脂肪酸を分解すると肝臓でのエネルギー産生はできますが、そのエネルギーを肝外組織に与えることができていませんでした。また、細胞質にはコレステロール合成系の酵素の1つとして細胞質HMG-CoA合成酵素がありました。進化の過程で細胞質HMG-CoA合成酵素の遺伝子が複製で2コピーになり、その1つがミトコンドリアに局在するようになってミトコンドリアHMG-CoA合成酵素(図ではHMGCS2)ができました。これによって脂肪酸ベータ酸化から産生されるエネルギー物質アセチル-CoAからHMG-CoAへの橋渡しを可能とし、元々あったロイシン代謝系のHMG-CoAリアーゼと結びつくことによって、アセト酢酸へと代謝されるケトン体産生系が誕生しました。これによってグルコースしか脳にエネルギーを供給できなかった状態からケトン体としてエネルギーを脳に供給できるようになり(もちろんほかの臓器にも)、哺乳動物はケトン体産生系を持つことで脳の発達に大きく貢献して来たと考えられています。飢餓に対して耐えることができるようになり、動物の進化において有利であったと考えられます。3日間の飢餓の後ではヒトの脳は25%のエネルギーはケトン体から得るとされ、さらに飢餓が進むとケトン体が脳の代謝の主要なエネルギー源となります。これは現在難治性のてんかんやGLUT1欠損症の治療にケトン食療法として応用されています。
6. 再び進化の過程でケトン体産生系を失った動物の系統が複数ある
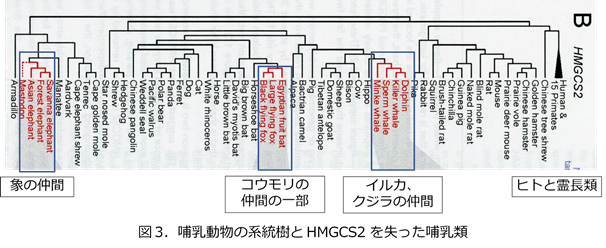
しかし、ケトン体産生系を進化の過程でふたたび失ってしまった哺乳動物種が複数あります。その多くはせっかく獲得したミトコンドリアHMG-CoA合成酵素の遺伝子が不活化されてしまったためです。ヒトにおいてはミトコンドリアHMG-CoA合成酵素欠損症として、飢餓時間が長くなった時に強い低血糖を起こしてしまう疾患ですが、これらの動物種はケトン体を産生できなくなった体質を優位に進化の中で選択して現在に至っております。これはケトン体代謝異常症を専門としている私にとっては驚くべき事実でした。どのような動物がケトン体産生系を失って進化してきているかといいますと、イルカやクジラの仲間、象の仲間、コウモリの一部の種類となります(図3)。これらの3系統はまったく系統樹上独立しており、動物進化の異なった場面で独立してケトン体産生系を失ったことになります。確かにクジラやイルカは飢餓状態になることはなさそうですよね?いつでも泳いで餌を取れそうです。象はどうでしょうか?餌に困ることはあまりないのかもしれません。これらのコウモリの仲間はやはり飢餓に弱いそうです。イルカやクジラの仲間、象の仲間ではこれらの脳容量の増大前にケトン産生系を失っており、ケトン産生系を失ってもこれらの種はきちんと脳を成長させていることは特記すべきことかもしれません。また、これらの哺乳動物ではケトン体産生系を失ったことが進化上優位になって進化してきたということは、失うことでメリットがあったはずなのですが、それは今のところ不明です。もしかしたらヒトおいても、ケトン体産生系を失うことで生命のリスクはあるわけですが、何らかのメリットもあるのかもしれません。
参考文献
上記ミトコンドリアHMG-CoA合成酵素の話は以下の論文から引用しました。
Jebb D and Hiller M: Recurrent loss of HMGCS2 shows that ketogenesis is not essential for the evolution of large mammalian brains. Elife. 2018 Oct 16;7. pii: e38906. doi: 10.7554/eLife.38906.
全文PDFは以下からダウンロードできます。